Iron limitation also affects the twilight zone
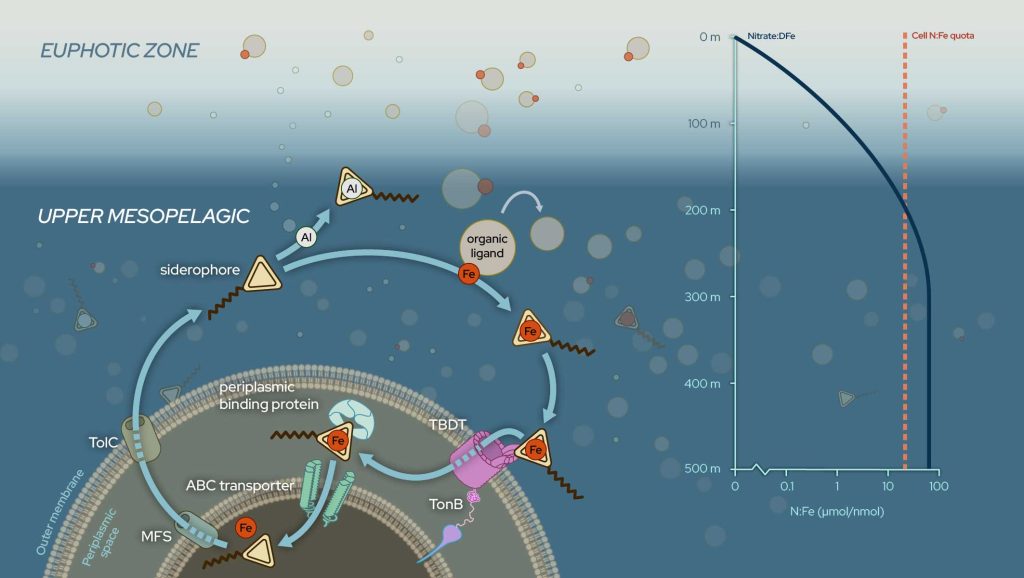
Iron limitation is known to affect the phytoplankton growth in the oceanic surface waters. Less know (but also less studied!) is its role in shaping microbial production in the mesopelagic layer, also called the twilight zone (200-500m below the surface). Siderophores are bacterial metabolites that convert iron bound to proteins or water-soluble compounds into a form accessible to microorganisms. Consequently, siderophores are biomarkers for microbial iron deficiency: the less iron is available, the more efficient the uptake must be.
Li and co-workers (2024, see reference below) established the distribution and uptake of siderophores along the GEOTRACES cruise GP15 (Pacific Meridional Transect). They reveal that concentrations are high not only in iron-limited surface waters but also in the twilight zone underlying the North and South Pacific subtropical gyres. They propose that such bacterial Fe deficiency owing to low iron availability also occurs in twilight zones of other large ocean basins, greatly expanding the region of the marine water column in which nutrients limit microbial metabolism.
Reference:
Li, J., Babcock-Adams, L., Boiteau, R. M., McIlvin, M. R., Manck, L. E., Sieber, M., Lanning, N. T., Bundy, R. M., Bian, X., Ștreangă, I.-M., Granzow, B. N., Church, M. J., Fitzsimmons, J. N., John, S. G., Conway, T. M., & Repeta, D. J. (2024). Microbial iron limitation in the ocean’s twilight zone. Nature, 633, 823–827. Access the paper: 10.1038/s41586-024-07905-z